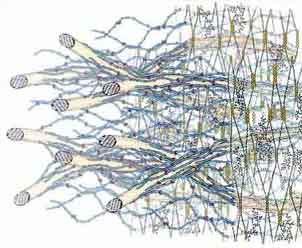
Arabidopsis and most angiosperms make a “Type
I” primary cell wall
For extensive review of the differences between grass species and all other
angiosperms with respect to polymer structure, architecture, and biosynthesis,
see McCann and Roberts (1991), Carpita and Gibeaut (1993), and Carpita (1996).
Briefly, “Type I” walls, which are found in dicots and the non-commelinoid
monocots, contain about equal amounts of cellulose and cross-linking xyloglucans
(XyGs), with various minor amounts of arabinoxylans, glucomannans, and galacto-glucomannans.
Except for the arabinosylated XyGs of Solanaceae, the general feature all
XyGs of Type I walls share is a repeating heptasaccharide unit structure subject
to further derivatization by additional galactosyl, fucosyl, and arabinosyl
units. XyGs occur in two distinct locations in the wall: binding tightly to
exposed faces of glucan chains in the cellulose microfibrils, and spanning
the distance between adjacent microfibrils or simply twining with other XyGs
to space and lock the microfibrils into place.
The cellulose-XyG framework of Type I walls is embedded in a pectin matrix that controls several physiological properties, such as wall porosity, charge density, and microfibril spacing (Willats et al. 2001). The two major pectins are homogalacturonans (HGAs) and rhamnogalacturonan I (RG I). Some HGAs and RGs are crosslinked by ester linkages to pectins or other polymers held more tightly in the wall matrix and can only be released from the wall by de-esterifying agents. Neutral polymers (arabinans or galactans) are pinned to one end to the RG I backbone, but extend into, and are highly mobile in, the wall pores. An unusual polysaccharide called rhamnogalacturonan II (RG II) has the richest diversity of sugars and linkage structures known. At some stages of cell development, hydrolases are released that trim these neutral polymers. Some Type I walls also contain several types of structural proteins that may interact with the pectin network. The various structural proteins could form intermolecular bridges with other proteins without necessarily binding to the polysaccharide components (Showalter 1993, Cassab 1998).
The cell walls of the Poales have different wall components
and architectures
Maize and other commelinoid monocots possess a different kind of primary wall,
a “Type II” wall (Carpita and Gibeaut 1993; Carpita 1996). They
contain cellulose microfibrils of the same structure as those of the Type
I wall, but glucuronoarabinoxylans (GAXs) are the principal polymers that
interlock the microfibrils. Unbranched GAXs can hydrogen bond to cellulose
or to each other. The attachment of arabinose and glucuronic acid side groups
to the xylan backbone of GAXs prevents the formation of hydrogen bonds, diminishing
the extent of cross-linking between two unbranched GAX chains or GAX to cellulose.
Type II walls have small amounts of XyG, but these XyGs contain neither arabinose
nor fucose. In general, grasses are pectin-poor; with the exception of the
lack of fucose, grass pectins are similar in structure to those of dicots.
When grass cells begin to elongate, they accumulate mixed-linked ß-glucans
in addition to GAX. This type of ß-glucan is unique to the Poales and
is a rare example of a cell expansion-specific polysaccharide (Carpita et
al. 2001).
Grasses, which have very little structural protein compared with dicots and non-commelinoid monocots, have extensive interconnecting networks of phenylpropanoids that form primarily when cells stop expanding (Iiyama et al. 1990). In the non-lignified walls the principal hydroxycinnamate is ferulic acid, whereas in the lignified walls both ferulic and p-coumaric acid are found. Although much less abundant, sinapic acid, 5-hydroxyferulic acid and caffeic acid have also been reported (Ohashi et al. 1987). Ferulic acid is esterfied to the C5 of arabinofuranose sidechains of arabinoxylans (Ishii 1997). Ferulic acid can dimerize, either via a photochemical reaction leading to cyclodimers (Ford and Hartley 1989), or via radical-mediated dimerization which gives rise to diferulatedehydrodimers (Ralph et al. 1994). Ferulate dimers are thought to crosslink different polysaccharide chains in the cell wall.
Carpita, N.C 1996. Structure and biogenesis of the cell walls of grasses. Ann. Rev. Plant Physiol. Plant Molec. Biol. 47, 445-476
Carpita, N.C., D. M. Gibeaut. 1993. Structural models of primary cell walls in flowering plants: consistency of molecular structure with the physical properties of the walls during growth. Plant J. 3, 1-30.
Carpita, N. C., M. Defernez, K. Findlay, B. Wells, D. A. Shoue, et al. 2001. Cell wall architecture of the elongating maize coleoptile. Plant Physiol. 127, 551-565.
Cassab, G. I. 1998. Plant cell wall proteins. Annu. Rev. Plant Physiol. Plant Mol. Biol. 49, 281-309
Ford, C. W., R. D. Hartley. 1989. GC/MS characterization of cyclodimers from p-coumaric acid and ferulic acids by photodimerization -- a possible factor influencing cell wall biodegradability. J. Sci. Food Agric. 46, 310-310.
Iiyama, K., T. B. T. Lam, B. A. Stone. 1990. Phenolic acid bridges between polysaccharides and lignin. Phytochemistry 29, 733-737.
Ishii, T. 1997. Structure and function of feruloylated polysaccharides. Plant Sci. 127, 111-127.
McCann, M. C., K. Roberts. 1991. Architecture of the primary cell wall. In: The Cytoskeletal Basis of Plant Growth and Form (C. W. Lloyd, ed.). London: Academic Press, pp. 109-129
Ohashi, H., E. Yamamoto, N. G. Lewis, G. H. N. Towers. 1987. 5-Hydroxyferulic acid in Zea mays and Hordeum vulgare cell-walls. Phytochemistry 26, 1915-1916.
Ralph, J., R. D. Hatfield, S. Quideau, R. F. Helm, J. H. Grabber, H.-J. G. Jung. 1994. Pathway of p-coumaric acid incorporation into maize lignin as revealed by NMR. J. Am. Chem. Soc. 116, 9448-9456.
Showalter, A. M. 1993. Structure and function of plant cell wall proteins. Plant Cell 5, 9-23
Willats, W. G. T., C. Orfila, G. Limberg, H. C. Buchholt, G. J. W. M. van Alebeek, et al. 2001. Modulation of the degree and pattern of methyl-esterification of pectic homogalacturonan in plant cell walls - Implications for pectin methyl esterase action, matrix properties, and cell adhesion. J. Biol. Chem. 276, 19404-19413.
Lignin is formed by specialized cell
Lignin is a complex, hydrophobic secondary cell wall polymer typically found
in tracheary elements and sclerenchyma that provides structural rigidity,
a water-proofing of certain cells, and a mechanical barrier against pests
and pathogens. The three most common monolignols are p-hydroxycoumaryl alcohol,
coniferyl alcohol and sinapyl alcohol, are synthesized via the shikimic and
phenylpropanoid pathways. Virtually all the genes that encode the enzymes
that function in monolignol synthesis have now been identified, including
one of the first steps, a 3’-hydroxylase involved in the synthesis of
caffeic acid form p-coumarate (Schoch et al. 2001), and one of the last steps,
a sinapyl alcohol dehydrogenase (Li et al. 2001). Polymerization into lignin
occurs via radical coupling in the wall. After polymerization lignin subunits
of these three monolignols are referred to as p-hydroxyphenyl, guaiacyl and
syringyl residues, respectively. Most dicot lignins are made of guaiacyl and
syringyl subunits. Grass lignins contain all three subunits, although the
proportion of p-hydroxyphenyl residues is typically less than 5% (Chen 1991,
Halpin et al. 1998). In grasses, ferulic acid is covalently bound to lignin
and is hypothesized to provide a nucleation site from which the lignin polymer
grows (Ralph et al. 1995). During the last 10 years major progress has been
made in the identification of the genes and enzymes involved in lignin biosynthesis
(for review, Whetten et al. 2001, Boudet 2000, Mellerowicz et al. 2001). Despite
this progress there is still a lack of detailed knowledge on variation in
lignin composition between species, and how a given plant species controls
tissue- and cell-specific variation in lignin subunit composition. Specifically,
the mechanisms behind monolignol transport and incorporation in the growing
lignin polymer need to be fully elucidated. A major question is whether monolignols
are coupled to each other in a random fashion (Hatfield and Vermerris, 2001),
or whether dirigent proteins are involved in this process (Davin and Lewis,
2000). Cell wall mutations have been identified in six stages of primary wall
biogenesis.
Boudet, A-M. 2000. Lignins and lignification: selected issues. Plant Physiol. Biochem. 38, 91-96.
Chen, C.-L. 1991. Lignins: occurrence in woody tissues, isolation, reactions, and structure. M. Lewin, and I. S. Goldstein (eds.). (New York:Marcel Dekker, Inc.), pp. 183-261.
Davin, L. B., N. G. Lewis. 2000. Dirigent proteins and dirigent sites explain the mystery of specificity of radical precursor coupling in lignan and lignin biosynthesis. Plant Phys. 123, 453-461.
Halpin, C., K. Holt, J. Chojecki, D. Oliver, B. Chabbert, et al. 1998. Brown-midrib maize (bm1) -- a mutation affecting the cinnamyl alcohol dehydrogenase gene. Plant J. 14, 545-553.
Hatfield, R., W. Vermerris. 2001. Lignin formation in plants: the dilemma of linkage specificity. Plant Physiol. 126, 1351-1357.
Li, L., X-F. Cheng, J. Leshkevich, T. Umezawa, S. A. Harding, V. L. Chiang. 2001. The last step of syringyl monolignol biosynthesis in angiosperms is regulated by a novel gene encoding sinapyl alcohol dehydrogenase. Plant Cell 13, 1567-1585.
Mellerowicz, E. J., M. Baucher, B. Sundberg, W. Bourjan. 2001. Unravelling cell wall formation in the woody dicot system. Plant Mol. Biol. 47, 239-274.
Ralph, J., J. G. Grabber, R. D. Hatfield. 1995. Lignin-ferulate cross-links in grasses: active incorporation of ferulate polysaccharide esters into ryegrass lignins. Carbohydrate Res. 275, 167-178.
Schoch, G., S. Goepfert, M. Morant, A. Hehn, D. Meyer, et al. 2001. CYP98A3 from Arabidopsis thaliana is a 3©ˆ-hydroxylase of phenolic esters, a missing link in the phenylpropanoid pathway. J. Biol. Chem. 276, 36566-36574.
Whetten, R. W., Y-H. Sun, Y. Zhang, R. Sederoff. 2001. Functional genomics and cell wall biosynthesis in loblolly pine. Plant Mol. Biol. 47, 275-291.
Back to top